









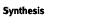
|
Faunal
Remains
by Robert J. Muir and Jonathan C. Driver
Introduction
1
In this chapter, we report on the analysis of faunal remains collected
during excavations at Yellow Jacket Pueblo (Site 5MT5) by the Crow Canyon
Archaeological Center. The assemblage includes all nonhuman bones and
teeth, antler, eggshell, and ossified cartilage found at the site. Both
modified and unmodified materials were analyzed and are reported on here.
2
The chapter begins with a brief discussion of the methods used to identify,
catalog, and quantify the faunal remains, followed by a description of
the animal taxa identified in the assemblage. Much of the chapter is given
to consideration of the taphonomic processes that might have influenced
the composition of the assemblage—that is, the various natural and
cultural processes related to deposition and preservation that likely
affected the kinds of remains that were recovered, as well as the condition
in which they were found. This discussion includes an assessment of the
natural and cultural modifications evident on the specimens and a reporting
of the skeletal representation of the most common taxa. Following the
discussion of taphonomy is an intrasite analysis of the spatial and chronological
distribution of the faunal remains, by architectural block. Finally, we
compare the faunal assemblage from Yellow Jacket Pueblo with the assemblages
from other Pueblo III sites in the region in an attempt to discern similarities
and differences in intrasite spatial patterning that might reflect economic
and/or social variation among communities.
Methods: Identification, Recording, and Quantification of Faunal Remains
3
All faunal remains collected during the excavations were analyzed by the
authors and cataloged using a standardized identification and recording
system developed by Jon Driver for Crow Canyon (Driver
1992*2). The following information was recorded for each specimen:
taxon, element, part of element, side, state of epiphyseal fusion, type
of breakage, modifications (cut marks, grinding, burning, weathering,
gnawing, etc.), length of fragment, and cortical thickness. Identifications
were made using comparative collections at Simon Fraser University, the
University of Puget Sound Museum of Zoology, and the Burke Memorial Museum
(Washington). We also used several osteological keys (including Gilbert
et al. 1981*1; Lawrence
1951*1; Olsen 1964*1,
1968*2; Schmid
1972*1) to assist us in sorting and making preliminary identifications.
Those interested in a more detailed description of identification and
cataloging procedures should consult additional sources (Driver
1992*1, 1992*2; Driver
et al. 1999*1).
4
Before identifying and cataloging the remains, we made a considerable
effort to reconstruct elements that had been broken during or after excavation.
We made no attempt to reconstruct elements that displayed old breaks—that
is, breaks that occurred before excavation. Fragments that were obviously
parts of the same bone were noted in the catalog. By preserving fragmentation
caused by cultural or natural taphonomic processes, we were able to view
the assemblage as it would have existed just before excavation. In the
subsequent analysis, each fragment or reconstructed element was counted
as a distinct specimen.
5
During analysis, a specimen was considered "identifiable" only if the
skeletal element could be positively determined. General element categories
such as "long bone" or "axial" were not used. All specimens that could
not be identified to a specific element were thus classified taxonomically
as "unidentifiable." This requirement ensures that analyses are not unduly
biased by the intuition and guesswork of individual analysts (Driver
1992*1).
6
All identifiable specimens were assigned to the most specific taxonomic
category possible, given the limitations of the available reference collections
and observable morphological variation. Bones were assigned to a species
or genus only when all other possibilities had been examined and ruled
out on the basis of morphology and size. Species-level identifications
were made only by direct comparison with modern skeletons. Many specimens
were assigned to more general taxonomic categories as defined by Driver
(1992*2).
7
Frequency data for the faunal remains from Yellow Jacket Pueblo are provided
as "number of identified specimens," or NISP counts (Grayson
1979*1). NISP counts represent the total number of specimens that
can be positively identified as belonging to a particular taxon. This
method has a number of potential problems (for a thorough discussion,
see Grayson [1979*1]). In
particular, NISP data will overrepresent taxa with (1) greater numbers
of elements (Klein and Cruz-Uribe
1984*1; Payne 1972*1);
(2) greater degrees of fragmentation (Grayson
1973*1, 1979*1; Thomas
1969*1; Watson 1979*1);
or (3) higher rates of recovery (Thomas
1969*1; Watson 1972*1).
Furthermore, NISP counts will produce artificially inflated sample sizes
(Watson 1979*1). NISP data
have been used here to allow direct comparison to faunal data produced
by other researchers, but they do not provide a particularly precise estimate
of taxonomic abundance.
Identified Taxa
8
Crow Canyon's test excavations resulted in the collection of 9,132 bone,
tooth, antler, and ossified cartilage specimens from the site. Of these,
3,939 could be identified to element and thus assigned to a specific taxonomic
category (Table 1). Approximately
200 bird eggshell fragments were collected from the site but were not
included in the analyses and discussion presented in this chapter.
9
Mammal remains dominate the identifiable assemblage, accounting for 69.0
percent of the specimens. Birds represent approximately one third (30.8
percent) of the identified remains, and trace amounts of fish and reptile
remains make up the balance of the assemblage. The identified specimens
represent a minimum of 40 mutually discrete taxonomic groups, including
at least 28 mammal and nine bird taxa (as well as one fish and two reptile
taxa). Additional taxonomic categories may be represented in the remains,
as a considerable number of specimens have been assigned to general categories
such as "medium carnivore" and "large bird." The majority of these remains
probably belong to taxa already identified within the assemblage. For
example, most specimens identified as "large bird" are probably the remains
of Meleagris gallopavo (turkey) or of one of the other large
bird species already identified within the assemblage. Similarly, remains
identified as "medium artiodactyl" are undoubtedly of Odocoileus
sp. (deer), Antilocapra americana (pronghorn antelope), or
Ovis canadensis (bighorn sheep). On the contrary, the wide variety
of Muridae (deer mice, voles, etc.) is difficult to separate osteologically,
and it is possible that species in addition to those named are represented
by these remains. Similarly, remains identified as "Sciuridae" (squirrels)
may represent a number of species not listed.
Mammals
10
The mammal remains from Yellow Jacket Pueblo include a wide variety of
taxa, although many are represented by only a few specimens (Table
2). The lagomorphs are most common, representing more than 54 percent
of the mammalian subassemblage. Cottontails (Sylvilagus sp.) are
extremely abundant, whereas jackrabbits (Lepus sp.) are represented
in much smaller numbers. Two species of cottontail may be represented:
Sylvilagus audubonii and Sylvilagus nutallii. No attempt
was made to distinguish between these species. The Lepus remains
were not assigned to a species, although snowshoe hare (Lepus americanus)
can be ruled out on the basis of size. No pikas (Ochotona sp.)
were positively identified among the remains.
11
A considerable quantity of rodent remains was recovered from the site,
representing approximately 24 percent of the mammalian specimens. Most
of the elements that were identified to genus and species were mandibles,
crania, teeth, innominates, and major long bones. Other rodent elements
have been identified only to the family level. Small rodents, including
deer mice and voles (Muridae), woodrats (Neotoma sp.), and pocket
gophers (Geomyidae), are most numerous. These animals are probably underrepresented,
given the potential for their very small bones to be lost or overlooked
during excavation. The larger rodents include rock squirrel (Spermophilus
variegatus), prairie dog (Cynomys sp.), porcupine (Erethizon
dorsatum), and beaver (Castor canadensis). Single specimens
representing chipmunk (Eutamias sp.) and Abert's squirrel (Sciurus
aberti) round out the rodent remains.
12
The order Carnivora is represented by at least five taxa and makes up
2.4 percent of the mammalian remains. Canids (fox, coyote, dog, and wolf)
are by far the most common family of carnivore represented; only one species,
domestic dog (C. familiaris), was positively identified. Other
carnivores are represented in smaller quantities, including marten (Martes
americana), long-tailed weasel (Mustela frenata), badger (Taxidea
taxus), and lynx (Lynx sp.). The lynx remains may be of bobcat
(L. rufus) or Canada lynx (L. canadensis), although the
latter is less probable, given this species' preference for heavily forested
environments and northern latitudes (Wooding
1982*1:130–132).
13
Artiodactyl remains account for less than 9 percent of the mammalian assemblage.
At least five species are represented: deer (Odocoileus sp.),
pronghorn antelope (Antilocapra americana), bighorn
sheep (Ovis canadensis), elk (Cervus elaphus), and domestic
cattle (Bos taurus). Most of the artiodactyl remains were identified
simply as "medium artiodactyl." Among those remains that could be identified
more precisely, deer (Odocoileus sp.)—representing either
mule deer (Odocoileus hemionus) or white-tailed deer (Odocoileus
virginianus)—are most abundant.
14
Finally, the orders Perissodactyla and Insectivora are each represented
by a single specimen: a horse (Equus caballus) radius and a shrew
(Soricidae) mandible, respectively.
Birds
15
Two taxa—Meleagris gallopavo (turkey) and "large bird"—dominate
the bird (Aves) subassemblage; together these make up more than 90 percent
of the bird remains (Table 3).
Many of the "large bird" bones are probably turkey. Birds such as owls,
ravens, falcons, and hawks may also be represented by the "large bird"
category, though probably only in very small proportions. In addition
to turkey, small quantities of other Galliformes (including quail and
sage grouse) were identified.
16
The order Columbiformes (pigeons and doves) is represented by two specimens,
or 0.16 percent of the bird subassemblage. Identifications to the family
or genus level were not possible for these remains.
17
Birds of prey, including members of the orders Falconiformes and Strigiformes,
are also present in the Yellow Jacket assemblage (1 percent). Among these,
the remains of hawk (Buteo sp.), falcon (Falco sp.),
and short- or long-eared owl (Asio sp.) were identified.
18
Passerine birds make up less than 1 percent of the bird subassemblage.
A single raven (Corvus sp.) specimen is conspicuous and easily
identified because of its relatively large size. The common raven (C.
corax), which is native to the area, is likely the species represented,
but the smaller Chihuahuan raven (C. cryptoleucus) could not
be excluded. The other Passeriformes remains are clearly those of smaller
species.
Reptiles and Fish
19
NISP data for the nine reptile and fish specimens recovered from Yellow
Jacket Pueblo are presented in Table
4. The reptile subassemblage contains both lizard and snake remains.
Four fragmentary fish bones were also identified. Little effort was made
to specifically identify any of these specimens because the comparative
collections used did not include a complete range of species.
Taphonomy
20
Taphonomy is the study of the natural and cultural processes that affect
the deposition and preservation of organic materials. Cultural behavior
associated with artifact manufacture, food preparation, and refuse disposal
will influence the distribution and composition of archaeological assemblages.
Once deposited on the ground, objects may be moved by natural mechanical
forces such as wind, water, or burrowing animals. Other natural agents
such as moisture, sunlight, and bacteria may erode or destroy materials.
Soil chemistry also affects preservation. Bones and other organic materials
are especially vulnerable to alteration, damage, or destruction as the
result of such factors. In this section, we examine evidence of taphonomic
processes that have affected the composition and condition of the Yellow
Jacket Pueblo faunal assemblage.
Weathering of Remains
21
Animal bones can be damaged or destroyed
by exposure to moisture, wind, and sun; the effects of such weathering
may have a substantial impact on the composition of a faunal assemblage.
The primary variable affecting the degree of bone weathering is the rate
of burial: bones that become buried more quickly will be less affected
by the natural elements. Bones will also weather differentially depending
on various natural attributes of the bones, including density, size, and
grease content. In addition, cultural practices such as processing, cooking,
and disposal may influence a bone's susceptibility to weathering. For
any of these reasons, bones of certain species may be more likely to suffer
from weathering and, thus, less likely to be recovered and/or identified
at an archaeological site.
22
In the Yellow Jacket Pueblo faunal assemblage, it is evident that some
taxa have been more severely affected by destructive weathering processes
than others (Table 5). In particular,
specimens from many of the medium and large mammalian taxa display evidence
of damage due to weathering. Weathering is also apparent on small percentages
of the "large bird," Meleagris gallopavo, and lagomorph remains.
Weathering is not apparent among the vast majority of small mammal remains,
and no medium or small bird remains show evidence of weathering. It is
perhaps most interesting that no small rodent remains appear weathered,
despite their relative abundance in the assemblage. The variability in
degree of weathering seen here may be due to both cultural and natural
processes. Whatever the cause, it does appear that the remains of some
taxa, particularly the medium and large mammals, are more likely to have
suffered from weathering and therefore may be underrepresented in the
assemblage.
Cultural vs. Natural Bone Accumulations
23
In general, the introduction of animal remains into archaeological sites
will occur as the result of one of four processes: (1) death of domesticated
animals raised on the site by humans; (2) killing of trapped or hunted
animals transported to the site by humans; (3) transport of dead animals
to the site by nonhuman predators or scavengers; or (4) natural death
of wild animals that lived and died on the site. Whereas the first two
processes relate directly to the human occupation of the site, the latter
two may be independent of human occupation and thus unrelated to cultural
activities; they may occur before, during, or after occupation of the
site.
24
Identification of cultural modification of medium and large mammal remains
can usually be made with confidence using discrete criteria such as breakage
patterns and the presence of cut marks; this assessment is more difficult
for the remains of smaller animals, which can be procured, processed,
cooked, and eaten with little modification to skeletal elements. It is
therefore impossible to assess how each individual specimen came to be
part of the assemblage. By considering the artificially and naturally
produced modifications observed on these remains, we hope to determine
the most probable taphonomic origin for the specimens of each taxon.
Culturally Modified Bone
25
Butchery and processing of animals can produce diagnostic modifications,
including cut marks, saw marks, or cut edges. Use of animal bones as tools
may also produce modifications such as evidence of grinding and polishing.
Remains that display such evidence are obviously indicative of human procurement
of the animals that they represent, although such markings are not likely
to occur on all, or even a majority, of culturally introduced specimens.
Evidence of burning, such as carbonization or calcinization, may be common
on culturally utilized taxa, but naturally deposited bones may, of course,
become charred as the result of wildfires or accidental house fires (Grayson
1988*1; Lyman 1988*1).
Spiral fractures are frequently considered indicative of human activity,
although these can also be produced by several natural agents, including
carnivores (Binford 1981*2),
natural traumatic injury (Lyman
1984*1), or trampling (Haynes
1983*1).
26
Based on evidence of cultural modification observed in the Yellow Jacket
Pueblo faunal assemblage (Table
6), a cultural origin for several specific taxa can be inferred with
considerable confidence. These include Lepus sp., Sylvilagus
sp., Castor canadensis, Canidae, Odocoileus sp., Buteo
sp., and Meleagris gallopavo. All display direct cultural modification
in the form of grinding, polishing, and cut marks. For some taxa, such
modification is extremely common. This is particularly true of the Odocoileus
sp. remains, of which 14.3 percent have been made into tools
or other artifacts. Numerous specimens in each of these taxa also display
burning or spiral fractures. The origins of the remaining taxa are uncertain,
as these groups display no definitive evidence of cultural modification.
Naturally Modified Bone
27
Bones from animals that were killed and/or consumed by predators and scavengers
may display several distinctive modifications. Most obvious are tooth
marks left by mammalian carnivores (Binford
1981*2), although the bones of very small animals may be totally consumed
by predators, such as owls, without extensive modification (Andrews
1990*1; Dodson and Wexlar
1979*1; Kusmer 1986*1).
Bones may also display spiral fractures (Binford
1981*2) or be extensively macerated (Korth
1979*1) as the result of carnivore predation. The ends of long bones
are often gnawed by carnivores and the diaphyses split to obtain marrow.
Licking of the ends of long bones by carnivores and other animals may
result in smooth, polished surfaces that closely resemble cultural modification
(Binford 1981*2; Haynes
1980*1). In such cases, crushed bone and tooth marks may be evident
elsewhere on the specimen. Acid etching of cortical bone may be evident
on bone fragments that have passed through a carnivore's digestive tract
(Andrews 1990*1:30). The
identification of specimens introduced to the site by carnivores is complicated
by the fact that the ancestral Pueblo are known to have kept domestic
dogs. Dogs are likely to have had access to discarded animal remains and
could have easily produced modifications indistinguishable from those
made by wild carnivores.
28
Table 7 presents the frequencies
of carnivore modification to faunal remains from Yellow Jacket Pueblo.
Carnivore modification is evident in nine taxonomic groups. It is significant
that carnivore modification is relatively common among several taxa (medium
artiodactyls, Odocoileus sp., and Meleagris gallopavo) that
are almost certainly of cultural origin at the site. This may indicate
the presence of domestic dogs. Unfortunately, these data do little to
clarify the origin of the other taxa.
29
Unmodified, predominantly complete bones should result from the natural
deaths of animals living on a site (Driver
1985*1:18). Burrowing animals may die in their burrows and should
display little bone damage from weathering or trampling. Animals that
are killed by humans and then processed for food or raw materials are
more likely to be represented by fractured specimens, particularly long
bones, which are commonly broken to extract marrow. Table
8 presents a comparison of complete and fragmented long bones of animals
that might have naturally inhabited Yellow Jacket Pueblo and died there
in burrows or dens. To mitigate biases due to element size, recovery rates,
and identifiability, only major long bones are considered here. Specifically,
the humerus, femur, and tibia have been selected for consideration because
of their relatively large size and taxonomically diagnostic characteristics.
However, it should be noted that differences in the sizes of the animals
in these taxa may introduce some biases into the comparisons presented.
For example, due to the extremely small size of the long bones of Muridae
(deer mice, voles, etc.), only complete specimens are likely to be recovered.
30
The data presented in Table 8
indicate that the Lepus and Sylvilagus long-bone remains
are predominantly fragmented specimens, whereas complete elements are
more common among the other taxa, particularly the small rodents. As mentioned
above, the frequencies may reflect, in part, the influence of animal size
on recovery rate, particularly for the smaller rodents; however, the differences
between the similarly proportioned Sciuridae and Sylvilagus remains
are unlikely to have resulted from factors associated with recovery rates
or identifiability. Thirty-three percent of the Sciuridae long bones are
complete, whereas less than 5 percent of the lagomorph long bones are
complete. The fragmentation data support the argument that the taphonomic
histories of the lagomorphs and rodents are considerably different. Specifically,
the abundance of complete rodent long bones is consistent with the suggestion
that these animals died naturally in their burrows. Conversely, the high
frequency of fragmentation among the lagomorph long bones lends further
support to the conclusion that these specimens are primarily a product
of cultural activity (as suggested by the evidence of cultural modification
presented above). Similar patterns have been documented at several other
sites in the central Mesa Verde region (Driver
et al. 1999*1; Muir 1999*2).
Modern Bone
31
Like most archaeological sites, Yellow Jacket Pueblo has been subject
to postdepositional disturbance by a variety of agents. Rodent burrowing
and other animal activities have undoubtedly been ongoing since establishment
of the site. It is also well known that Yellow Jacket Pueblo has been
heavily disturbed by artifact collectors, and it is possible that these
activities have resulted not only in mixing of deposits and removal of
artifacts, but also in introduction of materials to the site assemblage.
The remains of two taxa can be confidently assessed as intrusive to the
cultural deposits at Yellow Jacket Pueblo: cattle (Bos taurus)
and horse (Equus caballus). Neither of these species were present
in North America during the occupation of the site. The presence of these
specimens confirms that some mixing of modern and prehistoric deposits
has occurred.
Taphonomic Origins of the Faunal Remains
32
On the basis of the analyses and discussion presented above, we can draw
some conclusions regarding the taphonomic origin of the faunal remains
at Yellow Jacket Pueblo. Several taxa can be confidently assessed as being,
at least in part, the result of human activities. These include all those
taxa that display definite indications of cultural modification (that
is, cut marks, polishing, or grinding), specifically, Meleagris gallopavo,
Odocoileus sp., Canidae, Lepus, Sylvilagus, Castor
canadensis, and Buteo sp. It is also possible that a significant
number of specimens representing these taxa were introduced to the assemblage
naturally. This is most likely for those taxa, such as Sylvilagus,
that display only rare indications of cultural modification.
33
A second group of taxa, consisting of the small- and medium-size rodents,
displays characteristics that suggest that their presence is primarily
the result of the natural occurrence of these animals at the site. It
should be noted that numerous rodent species are mentioned ethnographically
as having been trapped and eaten by Pueblo peoples (Gnabasik
1981*1). However, no evidence of butchering was observed among the
many rodent remains recovered from Yellow Jacket Pueblo. Instead, the
rodent remains primarily include complete, unmodified specimens, which
is consistent with the interpretation that they represent animals that
died naturally in their burrows. The sciurids (squirrels) may be an exception.
A higher incidence of long-bone fragmentation than is typical for this
group and evidence of spiral fractures and burning suggest that the presence
of some of the sciurid remains may be the result of cultural activities.
34
The origins of the remains of the other taxa are uncertain. Most of these
taxa are represented by relatively few specimens, and therefore we have
little evidence of their collective taphonomic history.
Element Frequencies
35
It is generally expected that most animals will be represented archaeologically
by more-or-less complete skeletons, but in some instances cultural and/or
natural processes may influence the relative frequencies of particular
skeletal regions, elements, or element parts. For example, large game
may be represented at a habitation site by only those elements carried
there by hunters, or alternatively, at a kill site by only those elements
that are left behind (Perkins
and Daly 1968*1; White 1953*1).
Smaller animals are less likely to be affected by such differential transport
of parts, but they may become disarticulated and distributed throughout
a site as a result of butchering and processing. Consistent and repeated
cultural practices may cause specific portions of some taxa to be selectively
preserved, destroyed, or removed from the archaeological assemblage.
36
Natural agents can have similar effects on frequencies of skeletal parts.
Carnivores may selectively remove or destroy specific elements of some
species, thus creating assemblages that contain incongruent element compositions
(Binford 1981*2:214–216;
Marean et al. 1992*1). Rodents
may collect elements of a particular size range or density, resulting
in removal of these elements from a site or their preservation within
a burrow (Hoffman and Hays 1987*1).
Natural mechanical dispersal mechanisms such as colluvial and fluvial
forces may also selectively affect skeletal part frequencies. For example,
vigorous fluvial action will separate elements that float easily from
those that do not (Voorhies
1969*1). By analyzing skeletal part frequencies, we can assess the
degree to which individual taxa have been affected by such factors. Although
it is seldom possible to determine the precise cause of incongruent element
frequencies, we may identify the most probable or predominant influences.
37
For this analysis, the remains for each taxon are assigned to one of seven
skeletal regions: cranial, axial, pectoral girdle, forelimbs (further
categorized into upper and lower), innominate, hind limbs (further categorized
into upper and lower), and phalanges (Table
9). It should be noted that some specimens could not be assigned to
a specific region, primarily those identified simply as "metapodial" or
"sesamoid." Several patterns are immediately apparent. Predictably, regions
with fewer skeletal components (for example, the shoulder and pelvis)
have consistently lower specimen frequencies. Otherwise, most taxa with
substantial numbers of identified specimens (that is, more than 100) are
well represented in all skeletal regions. The small rodents present an
exception to this pattern, as no phalanges were identified for these taxa.
This is not surprising: these small elements are unlikely to be consistently
recovered during excavation, and, because of morphological similarities
across different families, little effort was made to precisely identify
phalanges of small mammals during analysis.
38
The family Geomyidae (pocket gophers) displays a clearly skewed element
distribution: elements of the cranial region far outnumber those of all
other skeletal regions. This phenomenon has been documented at other sites
in the Southwest by researchers who have argued that it represents evidence
of human procurement of these animals (Shaffer
1992*1). Although this is certainly possible, it seems more probable
that the frequencies reflect recovery and identification biases. The only
elements in the skeleton of a pocket gopher that are likely to be collected
consistently are the cranium and, perhaps, the larger long bones (humerus,
tibia, and femur) and innominate, although even these can fall through
¼-inch mesh. Axial elements and phalanges, if recovered, are unlikely
to be identified precisely and, consequently, will most often be classified
as small mammal or small rodent. This phenomenon of higher-than-expected
frequencies of cranial elements is also evident to a lesser extent among
the slightly larger woodrat (Neotoma) and squirrel (Sciuridae)
remains, where cranial and hind-limb elements predominate.
39
Axial elements (ribs, sternum, and vertebrae) are prominent among the
medium-artiodactyl remains. This may suggest differential transport of
carcass portions to the site. However, all other skeletal regions are
also well represented, so the common occurrence of axial elements probably
reflects a high degree of fragmentation of these remains, rather than
a significant human behavioral pattern.
40
Overall, the element frequencies at Yellow Jacket Pueblo suggest that
complete animal carcasses were deposited at the site. The observed
variability among element frequencies can be accounted for by commonly
recognized recovery, identification, and quantification biases. No strong
patterns indicative of selective transportation or distribution of specific
butchery units are evident for any of the taxa.
Intrasite Variation
41
The distribution of faunal remains at Yellow Jacket Pueblo is examined
here in an attempt to identify spatial and/or chronological patterning
within the site. The distributions of the identified taxa are presented
by architectural block in Table
10; the analysis presented here focuses on the three major taxa presumed
to be of cultural significance: turkey, lagomorphs, and artiodactyls.
Throughout these comparisons, the term "turkey" is used to refer to the
sum of all remains identified as either Meleagris gallopavo or
"large bird." Because of the relatively small sample sizes, detailed spatial
analyses of the less common taxa are not likely to reveal meaningful patterns.
Likewise, architectural blocks that yielded fewer than 50 specimens of
the three major taxa considered here are excluded from these analyses.
42
The absolute and relative frequencies of the three major taxa recovered
from each architectural block are presented in Table
11. Although considerable variability is apparent between the architectural
blocks, much of this can be attributed to the small samples recovered
from several areas (that is, Blocks 800, 1000, 1100, 2000, 2100, 2200,
2300, and 3300). When these blocks are excluded from consideration, general
consistency among the architectural blocks is apparent. Collectively,
turkey and lagomorph remains dominate the block assemblages, accounting
for more than 80 percent of the identified remains in all blocks. Most
of the block assemblages contain less than 11 percent artiodactyl remains,
with two exceptions: Block 2400 (14.4 percent) and Block 1200 (17.0 percent).
There is, however, considerable variation in the relative frequencies
of turkey and lagomorphs. Turkey bones typically make up 30 to 55 percent
of the major taxa, although Blocks 900 and 1200 stand out with values
of 23.7 percent and 19.3 percent, respectively. Lagomorph frequencies
range between 35.8 percent and 69.5 percent, with no clear clustering
of values. There is no obvious association between the relative frequencies
of these two species and the spatial organization of the blocks. No clusters
of blocks are apparent in which a particular taxon is either dominant
or underrepresented.
43
Some of this variability, however, may be explained by chronology. Differences
in the relative abundance of artiodactyl remains could be related to the
occupational history of the site. Those architectural blocks and associated
cultural deposits that are believed to have been first established during
the late Pueblo II period (see "Chronology")
consistently display relatively high frequencies of artiodactyl remains
(Figure 1). Specifically,
Blocks 100, 500, 700, 2400, and 3200 have artiodactyl frequencies ranging
between 6.5 percent and 14.4 percent of the major taxa. Artiodactyl remains
are consistently less common among those architectural blocks that are
believed to represent primarily mid- to late Pueblo III occupations. This
latter group includes Blocks 200, 300, 400, 600, 900, 1200, and 2500.
The one major exception to this pattern is Architectural Block 1200 (the
great tower complex), which has the greatest abundance of artiodactyl
remains in the tested areas of the site, despite being among the latest
occupied architectural units. That the great tower complex has a relatively
unusual faunal assemblage may attest to the specialized function of this
architectural unit. It is also notable that the bi-wall "tower kiva" within
this complex also displays a higher-than-average abundance of artiodactyl
remains, when compared with other mid- to late Pueblo III structures.
As is discussed below, similar patterns have been documented at Sand Canyon
Pueblo, another very large Pueblo III site in the region.
44
In contrast to the artiodactyl remains, the frequencies of lagomorph and
turkey bones do not display obvious spatial or chronological patterning.
The relative abundance of these taxa appears to vary sporadically among
blocks and through time and may simply reflect normal sampling variability
in conjunction with factors associated with taphonomy and quantification.
45
When compared across all tested blocks, the relative frequencies of the
three major taxa vary somewhat with context. Nonstructural contexts were
found to contain a relatively high abundance of turkey remains (42.1 percent)
when compared with contexts from structures (32.7 percent). Conversely,
lagomorph and artiodactyl remains are on average more abundant among structures:
artiodactyl remains represent 12.0 percent of the major taxa recovered
from structures and 6.8 percent among nonstructural contexts; lagomorphs
represent 55.3 percent among structure contexts and 51.0 percent among
nonstructural contexts. We have documented this pattern at other Pueblo
III sites in the region, most notably Sand Canyon Pueblo (Muir
1999*2) and Castle Rock Pueblo (Driver
2000*1), and believe it may relate to either storage/disposal practices
or natural taphonomic factors (see paragraph 50 ).
Intersite Variation
Yellow Jacket Pueblo Compared With Other Pueblo III Sites in the Central
Mesa Verde Region
46
The authors have analyzed the faunal assemblages of three other Pueblo
III sites in the region—Sand Canyon Pueblo (Muir
1999*2), Castle Rock Pueblo (Driver
2000*1), and Woods Canyon Pueblo (Driver
2002*3)—and the results of these analyses provide a basis for
intersite comparisons (Table 12).
These comparisons are presented in more detail elsewhere (Muir
and Driver 2002*2) and will only be briefly summarized and discussed
here.
47
Overall, the Pueblo III faunal assemblage from Yellow Jacket is most similar
to that from Sand Canyon Pueblo, both in terms of composition and distribution
of the remains. In particular, the relative frequency and distribution
of artiodactyl remains are remarkably similar at the two sites. While
Pueblo III sites in the central Mesa Verde region typically yield assemblages
dominated by turkey and lagomorphs, only Sand Canyon and Yellow Jacket
pueblos contain substantial quantities of artiodactyl remains (Muir
and Driver 2002*2). In both assemblages, artiodactyls make up more
than 8 percent of the major taxa. These remains are found clustered in
association with particular structures at each site. Much like the analyses
performed for Yellow Jacket Pueblo, spatial analyses of faunal remains
from Sand Canyon Pueblo (Muir
1999*2) reveal a marked concentration of artiodactyl remains among
deposits associated with towers and a more uniform distribution of major
taxa throughout the remainder of the site. Artiodactyl remains are also
relatively abundant among the D-shaped bi-wall structure at Sand Canyon
Pueblo (Muir and Driver 2002*2).
Lagomorph remains are distributed broadly throughout Sand Canyon Pueblo
and are generally more common in structures than in midden areas. Turkey
remains are also common in both midden and structure deposits throughout
most of the site, but are relatively scarce within towers and the bi-wall
structure.
48
The faunal assemblages from Woods Canyon and Castle Rock pueblos are also
similar to the assemblage from Yellow Jacket Pueblo, though there are
some notable differences. Artiodactyl remains are relatively scarce at
both sites; they represent only 3.19 percent of major taxa identified
at Castle Rock and less than 0.5 percent at Woods Canyon. Turkey remains
are predominant in the Woods Canyon assemblage, representing more than
73 percent of the major taxa, whereas lagomorphs represent only 26 percent.
Relative frequencies of these taxa from Castle Rock Pueblo are very similar
to those from Yellow Jacket, with lagomorphs most abundant (57 percent),
followed by turkey remains (39 percent).
49
Intrasite analysis of the Woods Canyon fauna (Driver
2002*3) revealed only minor variations in the spatial distribution
of turkey remains and relatively uniform distribution of the other taxa.
The rim complex at Woods Canyon Pueblo, which contains several towers
and a D-shaped structure, did not display marked concentrations of any
particular taxa but, rather, was characterized by relative frequencies
typical of the site as a whole. The only significant spatial patterning
evident among the remains from Woods Canyon was revealed in comparisons
between depositional contexts, where lagomorph remains were found to be
more commonly associated with structures than with midden or courtyard
areas. Similarly, at Castle Rock Pueblo the only significant source of
intrasite variation was seen in the lagomorph remains, which were again
found to be more common in deposits in structures (Driver
2000*1).
Discussion
50
The broad distribution of both lagomorph and turkey remains throughout
all four sites and their predominance in midden assemblages suggest that
these animals were used widely and commonly throughout the villages. Both
turkey and lagomorphs are documented as having been primary sources of
meat for many historic Pueblo peoples (Gnabasik
1981*1), and the distribution of these taxa appears to be consistent
with that of common daily food refuse (or domestic household refuse).
However, lagomorph remains are more prevalent in structure deposits than
they are in nonstructural and midden areas. This is in contrast to turkey
remains, which are less abundant in structures. As we have discussed elsewhere
(Driver 2000*1; Muir
1999*2; Muir and Driver
2002*2), these patterns may reflect a wide range of factors, including
consequences of gender roles, value, status, spiritual associations, abandonment
ritual, or domesticity. We believe, however, that two factors stand out
as particularly probable explanations. Differences in the storage of these
animals would account for the observed patterns. Specifically, lagomorphs
are documented ethnographically as being stored whole (complete with bones)
within roomblocks, while no such practice appears to be documented for
turkeys (Gnabasik 1981*1).
This may explain the greater relative abundance of lagomorph remains within
structures. Alternatively, natural introduction of lagomorph remains to
the structure contexts may be responsible for their apparent abundance.
Cottontails may find the crevices and shelters provided by collapsed stone
masonry to be ideal locations for dens and burrows. However, our taphonomic
analyses of the assemblages at all sites consistently suggest that the
presence of lagomorphs is primarily the result of cultural activities.
51
The predominance of lagomorph and turkey remains at Yellow Jacket Pueblo
is typical of Pueblo III sites in the central Mesa Verde region. Driver
(2002*4) has demonstrated that the archaeological faunal record displays
a long-term regional trend toward an increased dependence on turkey and
a corresponding decrease in the exploitation of large game from Basketmaker
II through Pueblo III times. Given this trend, the relatively high abundance
of artiodactyl remains found in association with particular structures
at Yellow Jacket and Sand Canyon pueblos is intriguing in that it may
provide insight into the social and economic organization of these large
villages.
52
Muir (1999*2) has argued
that the distribution of artiodactyl remains at Sand Canyon Pueblo suggests
that remains found among towers and other associated structures are related
to communal hunting activities; further, certain architectural blocks
acted as hunting or war society houses (or offices), where the spoils
of communal hunts were processed and stored. Alternatively, bones of artiodactyls
might have been disposed of in particular locations or structures, possibly
to protect them from scavengers. Such "respectful" treatment of the remains
of game animals was widespread in North America (Beaglehole
1936*1; Luckert 1975*1;
Tanner 1979*1; Underhill
1946*1).
53
Lipe and Ortman (2000*1)
have suggested that multiwall structures functioned as residences for
families who had considerable ceremonial and political influence within
the community. The abundance of artiodactyl remains associated with these
structures may indicate that the residents of bi-wall structures had preferential
access to wild game. As discussed above, no such correlations between
structure type and faunal distribution were documented at Woods Canyon
and Castle Rock pueblos. It appears that activities that resulted in the
creation of such concentrations at the larger sites did not occur at these
smaller pueblos. This may indicate fundamental differences, corresponding
to community size, in the nature and organization of hunting activities
or in the distribution of the spoils of such hunts (Muir
and Driver 2002*2).
Conclusions
54
The faunal assemblage from Yellow Jacket Pueblo is typical of faunal assemblages
from large late Pueblo II/Pueblo III sites in the central Mesa Verde region.
Turkey and lagomorphs are the predominant taxa represented in the assemblage,
whereas the remains of artiodactyls are less common and become increasingly
scarce through the Pueblo III period. This pattern has been documented
as part of a gradual, long-term (Basketmaker II–Pueblo III) decrease
in dependence on large game and a corresponding increase in dependence
on domesticated species, as well as on wild animals that frequented garden
plots as "pests" (Driver 2002*4).
Skeletal representation for most taxa suggests that the bones are the
remains of complete animals that were brought to the site for butchery,
processing, and consumption. The presence of rodent remains recovered
from the site appears to be primarily the result of natural taphonomic
processes rather than cultural activities.
55
The spatial distribution of remains within Yellow Jacket Pueblo is similar
to that observed at Sand Canyon Pueblo. The concentration of artiodactyl
remains within and around specific structures at these sites suggests
that the procurement of large game was a communal activity. The great
tower complex at Yellow Jacket Pueblo might have been a facility in which
communal hunts were organized and/or communally acquired game was redistributed.
Alternatively, the complex might have been the residence or office of
political or spiritual leaders within the community. A larger sample of
faunal materials from the great tower complex and similar structures at
other sites in the region would be valuable for evaluating these possibilities.
Acknowledgments
This research was conducted with the cooperation
and assistance of Crow Canyon Archaeological Center researchers and staff.
We would like to thank all of the individuals who have been involved in
the Yellow Jacket project, particularly Kristin Kuckelman, Donna Glowacki,
Scott Ortman, and Mark Varien. Thanks are also owed to the Department
of Archaeology at Simon Fraser University, which provided facilities and
support. This research has been funded in part by grants to Dr. Jon Driver
from the Social Sciences and Humanities Research Council of Canada and
Simon Fraser University.
References
cited | To
borrow, cite, or request permission
|

{kind=link}